

Simple and rapid design of genetically engineered rodent models.
Customized mouse and rat model generation via the nuclease-based system CRISPR/Cas9 outcompetes traditional methods in terms of time. Additionally, CRISPRs can be designed to target virtually all genes in a eukaryotic genome, just by synthesis of short sequence specific RNAs.
Like TALENs, the CRISPR/Cas9 system (for review see Doudna and Charpentier, 2014 and Hsu et al., 2014 ) is used to introduce double strand breaks into a genome in a sequence specific manner. Whereas TALENs use a combination of hard-to-assemble protein domains recognizing the specific DNA sequence, the CRISPR/Cas9 system makes use of easily synthesized small RNA molecules that are applicable to a wider veriety of target sequences. These small RNAs, originally derived from Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) in bacteria, target the nuclease Cas9 to the specific genomic region and introduce double strand breaks (Jinek et al., 2012 ).
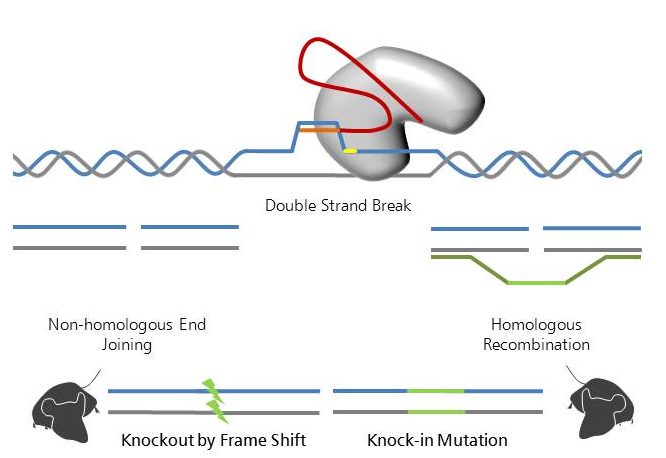
As compared to TALENs and other nuclease based techniques, CRISPR/Cas9 is the perfect system in terms of cutting efficiency. However, as the higher efficiency may lead to a higher number of off-target cutting for CRISPR/Cas9, a thorough analysis of off-targets is highly recommended.
CRISPR/Cas9 rodent model generation possibilities at PolyGene:
At PolyGene, CRISPR-based knockouts have all but replaced the traditional way of generating constitutive knockout mouse models (as well as knockout rat models).
For CRISPR/Cas9 microinjection into the oocyte, cf. Wang et al., 2013, Parikh et al., 2015, Paquet et al., 2016 for examples in mouse.

Larger insertions are more challenging and require complex strategies, e.g. when requiring coinjection of upstream and downstream guide RNAs, or targeting less permissive loci. In this instances, we will elaborate a careful alternative locus targeting strategy with the costumer.
For larger insertions, some groups have reported good efficiency for some “easy to targeted” loci like Rosa26 (Chu et al., 2016 ), but other genes have proven to be more difficult to target with homologous recombination in oocytes.
